|
# E8 x$ m( \% D! p
本期讲述海洋巨型病毒如何影响地球化学循环。 u5 [* ^+ M, _, r" P- O; S
巨型病毒为海洋捕食型单细胞提供视紫红质光合系统 # N0 A, J2 [; v
A distinct lineage of giant viruses brings a rhodopsin photosystem to unicellular marine predators
8 q# {9 y% c+ C1 ~ 作者:Needham DM, Yoshizawa S, Hosaka T, Poirier C等 / {& o% K- s U5 M
期刊:PNAS
4 Z, B7 h; L, A; x% Y J 时间:2019.10 , F4 O1 c$ F6 O
影响因子:9.58 / n! D% t; I: t% H. B8 k/ `
一、研究背景
$ ]& h/ l1 n# B& B: ? 病毒正被日益认为是海洋碳循环的关键参与者之一,特别是与光合作用相关的具有辅助代谢基因的病毒。过去已有报道发现拥有巨型基因组的病毒(>300 kb)可感染真核生物,即现在所说的巨病毒,过往认为巨病毒主要编码对细胞生命不起作用的蛋白,而目前发现它们还会编码辅助代谢蛋白,对感染宿主和改变宿主代谢过程中起到了重要的作用,从而影响了海洋生化循环。由于经典的病毒分离方法依赖于宿主培养后回收,而大部分的海洋原生动物(宿主)仍没有实现实验室培养,因此阻碍海洋病毒的研究发展。在此情况下,现在可以借助高通量测序技术在避免繁琐的实验室培养的同时获得样本中基因组水平的信息,尤其在降低样本多样性的条件下效果可明显提高,根据此思路,作者整合多种实验方法和技术,使用单细胞宏基因组技术获得未经培养的海洋巨病毒基因组,通过序列分析确定编码视紫红质的能力,并探讨了其全球海洋圈分布情况。
+ `& @4 w( j, V7 O. g6 z$ e 二、实验结果 : U. A- d' v) k" Q" ?5 m. ]
北太平洋捕食型原生生物及病毒的分离
, s& A9 [1 m8 Q+ y8 n 作者为了海洋中未被培养的异养型原生生物,使用酸性染料对样本中的细胞进行然而后再流式细胞仪(FACS)上分选原生生物和原核生物,额外再加上排斥光合有机体的筛选门仅保留异养型原生生物,最终成功地回收了鞭虫类单细胞生物。针对单个细胞的DNA的18S的V4区域进行扩增与测序分析发现其中有99%的序列注释为鞭毛虫,仅有1%的序列注释为寄生虫,推测此鞭毛虫中可能含有病毒病对此进行单细胞宏基因组测序与分析,成功组装一个875 kb的病毒基因组(ChoanoV1),是迄今为止通过测序获得最大的海洋巨病毒基因组。ChoanoV1的GC含量只有22%,远低于常规认知的巨病毒的GC含量(64%左右),其基因组预测能编码862种蛋白,可将其归纳为NCLDV(核质巨DNA病毒)。
# c; |( D. D" P7 Z8 y" r 在单细胞鞭毛虫中检测到巨病毒反映了几种可能的互作关系:1、该病毒感染了单细胞鞭毛虫并进行大量复制;2、该病毒作为猎物被单细胞鞭毛虫捕食;3、单细胞鞭毛虫捕食了被该病毒感染的细菌。目前有众多的证据支持第一种互作关系,如该病毒的平均测序深度达到215±157×,表明在宿主内高度复制,而在宿主中检测到的细菌序列深度远没有达到此水平,并且组装序列的N50也短于该病毒,说明病毒与宿主的直接关联性强。 6 Y, N& F! ^& f9 ~6 L, w) }4 n' x; [
作者在北太平洋东部离岸200公里处采集样本,并且通过密度梯度离心富集低GC含量的DNA进行测序与分析,组装得到ChoanoV2,其中89%预测具有编码能力的基因与ChoanoV1是一致的,说明作者的方法有助于研究病毒的进化、功能和代谢机制,具有一定的普适性。 - t/ h/ C' @) |
NCLDV巨病毒的进化分析
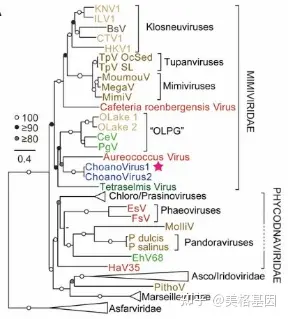
: i* u0 a0 `. ^ 初步分析表明,约有20%的ChoanoV1的预测蛋白与NCLDV进行Blast发现高关联性,而在ChoanoV2的比例是23%,但由于海洋病毒及原生生物的基因组信息缺乏,无法进行基因水平转移等研究。为了从另一个角度认识NCLDV的进化关系,借鉴植物遗传学的研究思路将NCLDV基因组核心蛋白进行构树,进化树表示ChoanoVirus属于Mimiviridae,并且形成一个独立的分支(图1),说明其独特性和普遍性。 0 c4 q- l. D7 P/ z
图1. NCLDV核心蛋白进化树ChoanoVirus辅助代谢基因对地球生化循环的意义
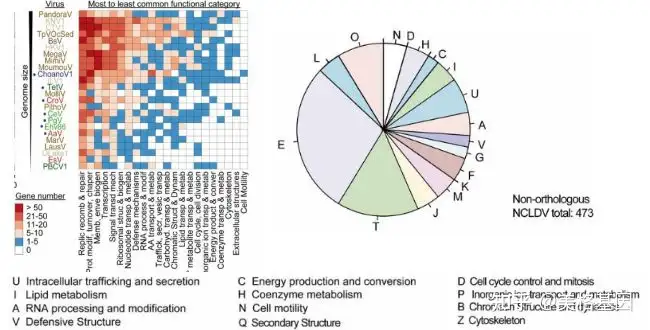
r! L) L! A! R7 }$ k5 l. Q! [; _ AMGs是由病毒携带的宿主衍生基因,与病毒的复制无关,但是能增强受感染的宿主的细胞功能,例如海洋噬菌体能在增强蓝藻光合作用能力。而ChoanoVirus的代谢潜能分析表明,预测能编码与光解酶、信号转导、DNA复制、细胞壁和翻译后修饰等相关的多种蛋白,并且还具有其他NCLDV所没有的特殊基因(图2)。特别是的ChoanoVirus能编码22种tRNA,而tRNA的数量似乎与基因组大小成正比,并且ChoanoVirus的tRNA数量与氨基酸使用量也是成正比,说明这些tRNA还是优先为ChoanoVirus的翻译过程服务,而不是为宿主的翻译过程服务。
& y: v( P+ ]* K0 |/ b( E$ | 图2. ChoanoVirus辅助代谢基因的注释结果展示 病毒视紫红质的序列特征和全球分布
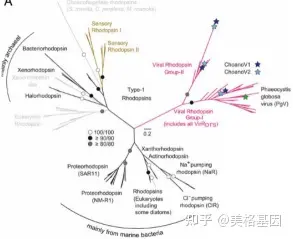
7 m8 ?3 }* ~; i/ s' } 视紫红质是不可或缺的膜蛋白,可结合发色团来感知阳光,而在ChoanoVirus基因组中鉴定出3种不同的视紫红质基因。系统发育树分析表明,ChoanoVirus的视紫红质可分成2大类型,表现出不同的进化关系(图3),作者将这两大类命名为:VirR-I与VirR-II。作者再结合拓扑分析推测,视紫红质并非源自于现存其他物种,而是存在于远古病毒并随着感染入侵和扩展到不同的宿主中。
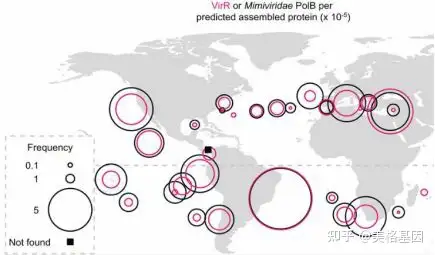
9 M! k5 N4 ], J7 X7 q; ] 图3. 视紫红质系统进化树目前视紫红质在全球的分布情况仍不清楚,因此作者对全球的VirR库进行搜索,在37个地点中得到相关序列信息,分析发现与海洋原核视紫红质吸收多种波长的光不同,超过99%环境中的视紫红质集中在于吸收绿光。并且在这37个地点中分发现视紫红质与Mimiviridae的相关性达到0.7,说明视紫红质基因是能接受阳光照射的海洋巨病毒基因组的常见组成部分。
* B+ L: @- D* y( ?* {9 ^ 图4. 视紫红质与巨病毒的全球分布情况三、亮点 , I+ y9 o: Z: G6 _! Y
1、病毒分选思路较为特别,富集目标清晰;
/ U" c( g/ v- A+ c7 z$ [5 d 2、对于病毒基因组序列挖掘思路清晰; ; c! [, [8 ]; D1 O4 p1 X
3、文章角度从病毒分离和病毒序列分析升华到地球化学循环,提升文章档次。 7 R; l/ y. d+ @1 R9 ?' e5 X0 J& t. y6 h
您可能还喜欢:
0 [7 p" b; @' `6 j7 ^5 B) [
/ A( c4 t$ z' b 2 E. J$ O% W9 x2 ^( A0 p* |
& w( B8 G( E/ \. j
+ [4 S0 u+ w# Z. b& a
+ f+ m2 I( C2 h K2 }6 m3 q6 |" m5 B' @( V
|